Om den irreduserbare kompleksiteten til spermceller
Jonathan McLatchie; 30. juni 2023
Oversatt herfra
Bilde 1. Spermceller
Menneskelig reproduksjon er kanskje det mest typiske eksempelet på teleologi i biologi. Prosessen der et befruktet egg utvikler seg til et spedbarn i løpet av ni måneder avslører utsøkt ingeniørkunst og genialt design. Før denne intrikate prosessen i det hele tatt kan begynne, er det behov for en sædcelle å smelte sammen med et egg - hver bærer, når det gjelder mennesker, 23 kromosomer. Denne utrolige bragden bærer de umiskjennelige kjennetegnene på bevisst hensikt og framsyn. Du kan se en animasjon av denne bemerkelsesverdige prosessen her:
Video: befruktning
Her vil jeg fokusere på designkarakteristikkene til sædceller. I et påfølgende innlegg vil jeg diskutere designfunksjonene til sædvæsken, og sædkapasitet. Sædceller består av tre komponenter - hodet, midtstykket og flagellen - og hundrevis av millioner av dem inneholdes i sædvæsken som slippes ut i livmorhalsen gjennom ejakulasjon under samleie. Ved hver utløsning frigjør hannen mellom to hundre og fem hundre millioner sædceller (omtrent 100 millioner per milliliter sæd). Hver av disse tre komponentene, og sædvæsken, er avgjørende for sædcellens oppdrag med å smelte sammen med et egg for å danne en zygote (et befruktet egg). La oss vurdere hver og en etter tur.
Hodet
Hodet bærer tett kveilede kromatinfibre, som inneholder det haploide genomet - totalt halvparten av det genetiske materialet som vil arves av neste generasjon (den andre halvparten vil komme fra morens eggcelle). Den tette innpakningen av DNAet tjener til å minimere volumet for transport.
På spissen av sædhodet er en membranøs organell, kalt akrosomet, som inneholder ulike hydrolytiske enzymer. Når disse skilles ut, fordøyer de eggcellemembranen, og letter derved penetrasjon av egget. Uten akrosomet vil sædcellen ikke være i stand til å trenge gjennom eggcellemembranen for å befrukte egget. I følge en gjennomgangsartikkel publisert i Frontiers in Cell and Developmental Biology:
"Enhver strukturell eller funksjonell akrosomal abnormitet kan svekke spermfusjon og til slutt resultere i infertilitet. Studier har dessuten vist at intra-cytoplasmatisk inseminasjon med sæd som inneholder akrosomale abnormiteter, ikke førte til vellykket befruktning, selv i fravær av befruktningsbarrierer, fordi oocytten ikke var i stand til å aktiveres effektivt .. Akrosomet er derfor uunnværlig for befruktning." (1)
Når en sædcelle når i nærheten av egget, gjennomgår den en rekke molekylære interaksjoner med zona pellucida, som er en spesialisert ekstracellulær matrise som omgir egget. Spesifikke reseptorer på spermens plasmamembran, som sperm-adhesiner eller -integriner, gjenkjenner og binder seg til tilsvarende ligander på zona pellucida. Denne bindingen utløser 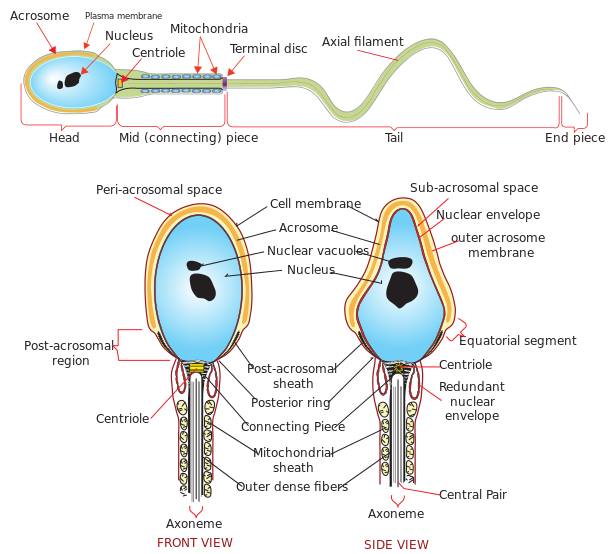 aktivering av signalveier i sædcellene. Binding av spermreseptorene til zona pellucida ligander fører til en tilstrømning av kalsiumioner (Ca2+) inn i sædcellen. Denne kalsiumtilstrømningen formidles vanligvis av ionekanaler eller reseptorer på sædcellens plasmamembran, som aktiveres ved ligand-reseptorbinding. Økningen i intracellulære kalsiumnivåer setter i gang en signalkaskade i sædcellen. Kalsiumioner fungerer som andre-budbringere, og utløser aktivering av forskjellige nedstrøms signalmolekyler og enzymer, inkludert proteinkinaser. Som et resultat av den kalsiummedierte signalkaskaden gjennomgår akrosomet eksocytose. Membranen som omgir akrosomet smelter sammen med sædcellens plasmamembran, og forårsaker frigjøring av det akrosomale innholdet, inkludert enzymer som hyaluronidase og acrosin. Enzymene som frigjøres fra akrosomet hjelper til med å bryte ned glykoproteinmatrisen til zona pellucida, slik at sædcellene kan trenge inn og nå eggets plasmamembran. Det akrosomale innholdet hjelper til med å bryte ned de beskyttende lagene som omgir egget, og letter sammensmeltingen av sæd- og eggmembranene.
aktivering av signalveier i sædcellene. Binding av spermreseptorene til zona pellucida ligander fører til en tilstrømning av kalsiumioner (Ca2+) inn i sædcellen. Denne kalsiumtilstrømningen formidles vanligvis av ionekanaler eller reseptorer på sædcellens plasmamembran, som aktiveres ved ligand-reseptorbinding. Økningen i intracellulære kalsiumnivåer setter i gang en signalkaskade i sædcellen. Kalsiumioner fungerer som andre-budbringere, og utløser aktivering av forskjellige nedstrøms signalmolekyler og enzymer, inkludert proteinkinaser. Som et resultat av den kalsiummedierte signalkaskaden gjennomgår akrosomet eksocytose. Membranen som omgir akrosomet smelter sammen med sædcellens plasmamembran, og forårsaker frigjøring av det akrosomale innholdet, inkludert enzymer som hyaluronidase og acrosin. Enzymene som frigjøres fra akrosomet hjelper til med å bryte ned glykoproteinmatrisen til zona pellucida, slik at sædcellene kan trenge inn og nå eggets plasmamembran. Det akrosomale innholdet hjelper til med å bryte ned de beskyttende lagene som omgir egget, og letter sammensmeltingen av sæd- og eggmembranene.
Bilde 2. Spermceller -skjematisk, i lengde og tverrsnitt
Dannelsen av selve akrosomet er delt inn i fire stadier. Det første stadiet, "Golgi-fasen", er avhengig av Golgi-apparatet, som produserer og pakker proteinene og enzymene som trengs for akrosomdannelse. Disse proteinene blir deretter transportert inn i den utviklende akrosomvesikkelen. I den andre fasen, "hettefasen", smelter den Golgi-avledede vesikkelen (kjent som den proakrosomale vesikkelen) sammen med den fremre delen av kjernen, og danner en hettelignende struktur over kjernen. Fusjonen av vesikkelen med kjernen formidles av membran-handelsprosesser. Den proakrosomale vesikkelen inneholder enzymer, glykoproteiner og andre komponenter som er essensielle for akrosommodning. I den tredje fasen, "akrosomfasen", gjennomgår den hettelignende strukturen en rekke strukturelle endringer, som fører til dannelsen av akrosomet. Den proakrosomale vesikkelen flater ut og forlenges, og sprer seg over den fremre delen av kjernen. De Golgi-avledede enzymene modifiserer proteinene som er tilstede i den proakrosomale vesikkelen, og omdanner dem til deres aktive former. Den akrosomale membranen gjennomgår også endringer, og blir spesialisert for akrosomets funksjoner. I den siste fasen, "modningsfasen", gjennomgår akrosomet ytterligere modifikasjoner og modning. Enzymer i akrosomet blir fullstendig aktivert og den akrosomale matrisen gjennomgår endringer, og blir mer kondensert. Det akrosomale granulatet, som er den sentrale delen av akrosomet, blir svært elektrontett på grunn av akkumulering av enzymer og proteiner. Det modne akrosomet er nå klart for sin rolle i befruktningen. For en mer detaljert beskrivelse av denne utrolige prosessen, henviser jeg leserne til en gjennomgangsartikkel om "Mechanism of Acrosome Biogenesis in Mammals." (2)
Mellomstykket
Midtstykket består av en sentral filamentøs kjerne, rundt denne er det mange strategisk plasserte mitokondrier som syntetiserer energimolekylet adenosintrifosfat (ATP). Kompleksiteten og utformingen av energi-generering i mitokondriene - inkludert prosessene med glykolyse, sitronsyresyklusen (eller Krebs), elektrontransportkjeden og oksidativ fosforylering - kan være en egen serie med artikler, men dette er et tema for en annen gang. For en god introduksjon til de fenomenale prosessene i mitokondriene, her er tre animasjoner fra Harvard University som bringer denne fascinerende organellen til live:
"Mitokondrier: Cellens kraftverk" -lenke
"Elektrontransportkjede" -lenke
"ATP-syntase i aksjon" -lenke
ATP generert av mitokondriene gir energi til flagellens kraftslag, og driver dens reise gjennom den kvinnelige livmorhalsen, livmoren og livmorrørene. Som sådan er den midterste delen av sædcellen helt avgjørende for dens funksjon å svømme gjennom den kvinnelige livmoren og egglederen for å befrukte egget hennes. Uten midtstykket og dets mitokondrier er sædcellene fullstendig ubevegelige.
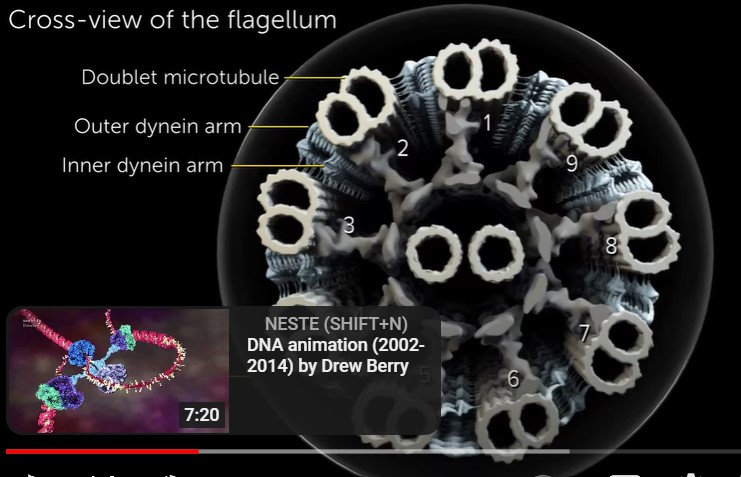 Flagellen
Flagellen
I motsetning til en bakteriell flagell (som roterer som en motor), slår en spermflagell med en pisklignende bevegelse for å produsere bevegelighet. Hvordan fungerer flagellen? I 2018 belyste Jianfeng Lin og Daniela Nicastro mekanismen for flagellarmotilitet.(3) Dataene deres indikerte at "bøyning ble generert av den asymmetriske fordelingen av dyneinaktivitet på motsatte sider av flagellumet"(4) (dyneiner er ATP-drevne molekylmotorer som "går" langs mikrotubuli mot minusenden -lenke ). Resultatene deres avslørte også at vekslende flagellbøyning oppstår på grunn av "en 'bryter-inhibering'-mekanisme der kraftubalanse genereres ved å hemme .. dyneiner på vekslende sider av flagellumet."(5) Med andre ord fører regulatoriske signaler til hemming av dynein. motorer på den ene siden av flagellen. I mellomtiden, på den andre siden, går dyneinene langs mikrotubuli. Flagellen bøyer seg i én retning på grunn av molekylære linkere som motstår denne glidningen. Flagellarbøyningen veksler ved gjentatte ganger å bytte siden av dynein-hemming. Se her for en animasjon som viser hvordan dette er tenkt å fungere -lenke.
Bilde 3. Sperm-flagell tversnitt
Det sier seg selv at uten flagellen er sædcellen fullstendig immobil og har ingen sjanse til å befrukte egget.
Så langt har vi vurdert den irreduserbare kompleksiteten til komponentene i en sædcelle. I en påfølgende artikkel skal vi vurdere designtrekkene til sædvæsken og prosessen med spermkapasitet som finner sted i den kvinnelige livmor-kanalen.
Fortsettelse -her.
Oversettelse via google ovesetter og bilder, ved Asbjørn E. Lund